Contributors
Paul G. Thomson1,2
Dion M.F. Frampton3
Lesley A. Clementson3
Chari B. Pattiaratchi1,2
1 Oceans Graduate School, The University of Western Australia, Crawley, WA, Australia
2 UWA Oceans Institute, The University of Western Australia, Crawley, WA, Australia
3 CSIRO, Oceans and Atmosphere, Hobart, Tasmania, 7000, Australia
Key Information
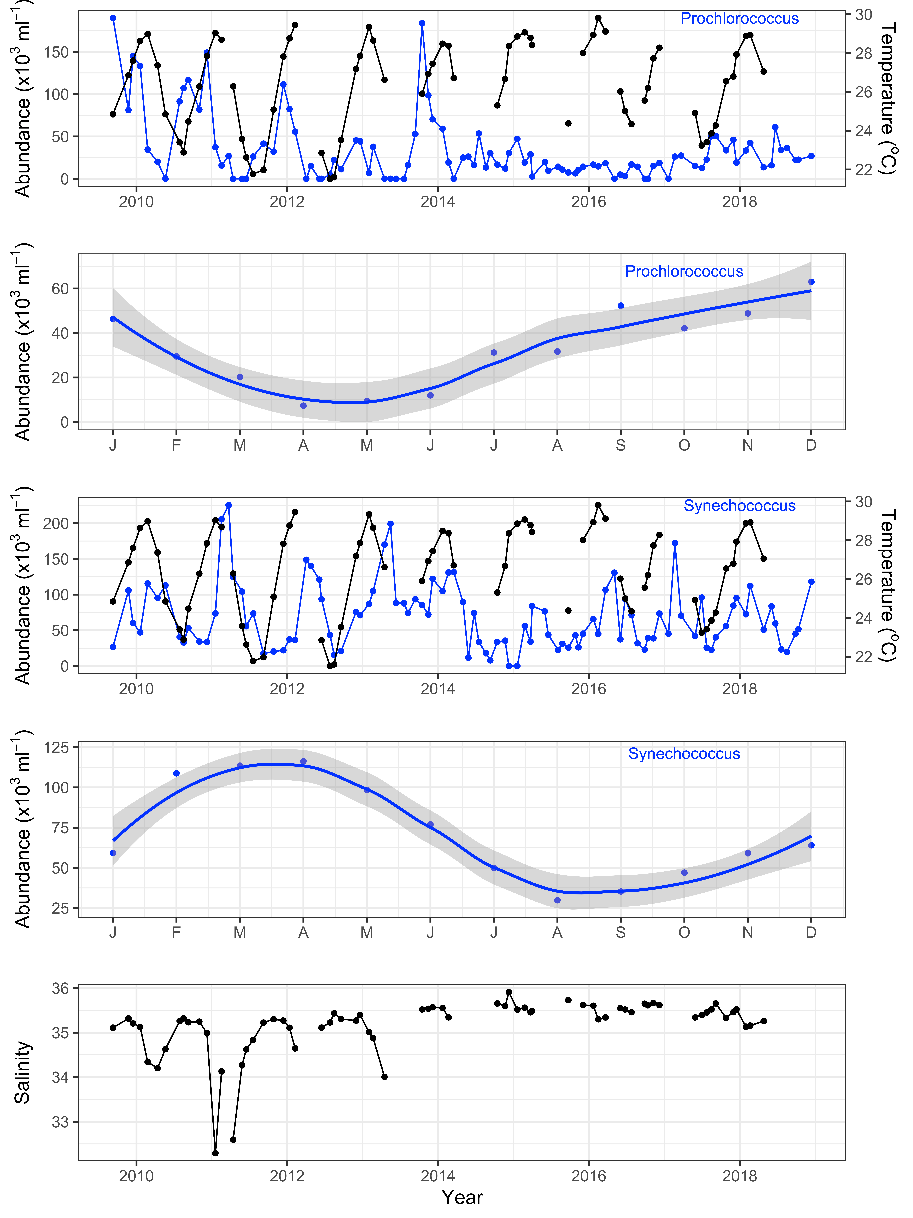
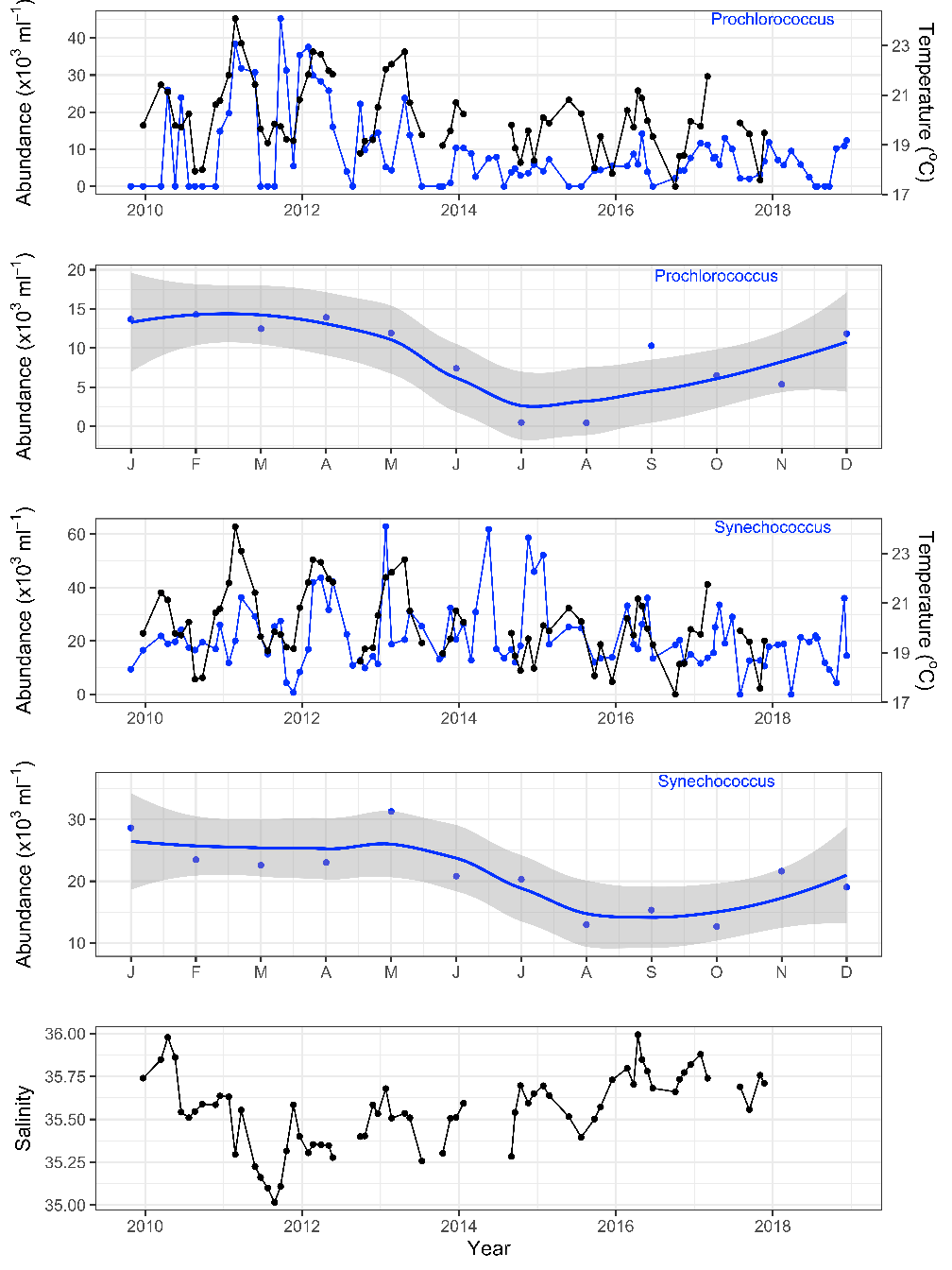
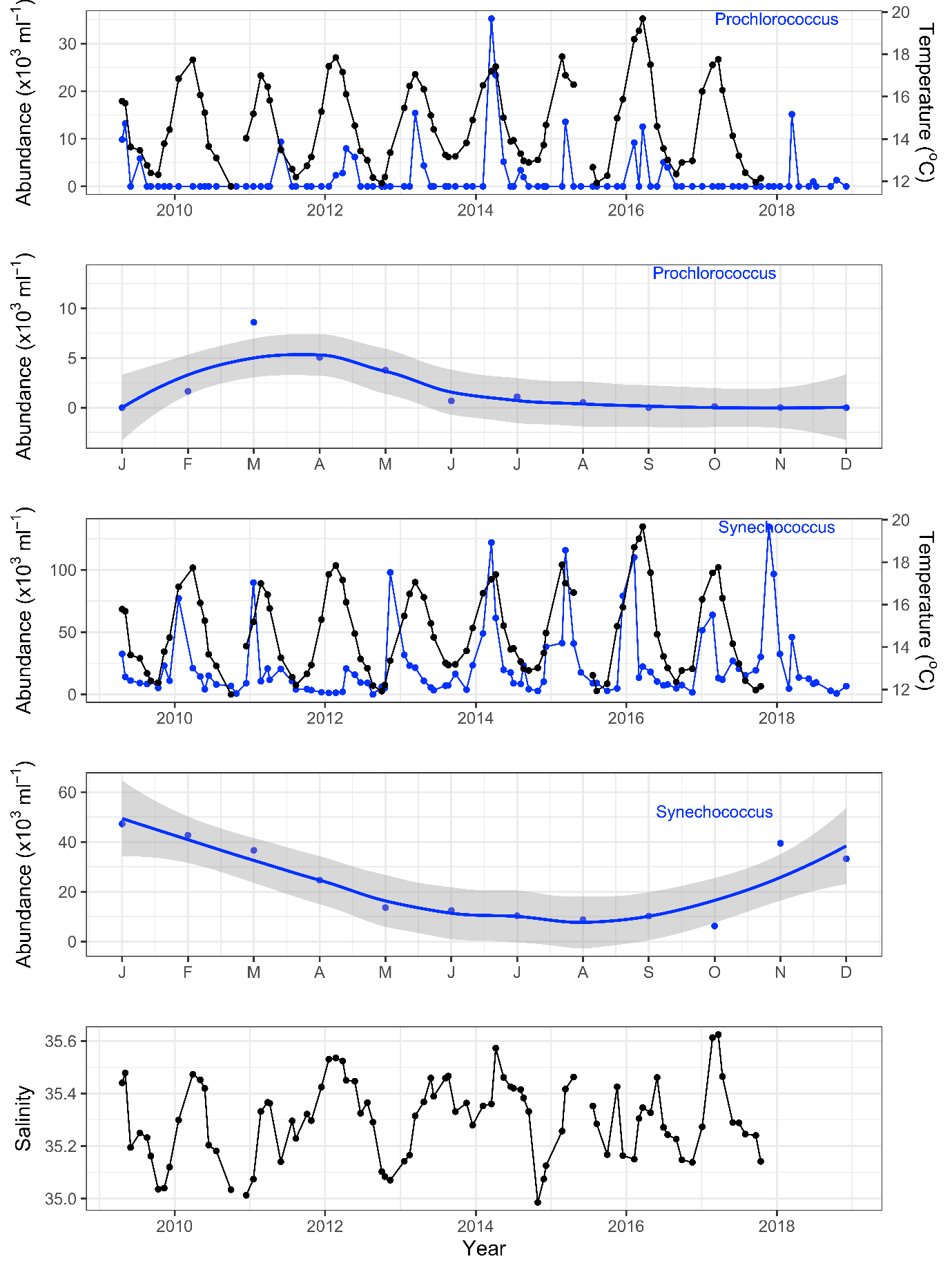
Strengthening boundary currents and episodic marine heatwaves are carrying tropical and subtropical picophytoplankton species such as Prochlorococcus and Synechococcus far into temperate southern waters around Tasmania and South West Australia, with implications for both people and ecosystems. As biomarkers for warmer currents, the picophytoplankton are sensitive indicators of tropical/subtropical microbial communities and thus may 1) afford us a glimpse of future microbial communities in our warming southern oceans and 2), help us model the spread of viral diseases, pathogens and other microbes along our coastlines. Using sensitive biomarkers such as the picophytoplankton will help us understand changes in marine microbial communities at the base of the marine food chain and subsequent impacts on the environment and marine industry such as aquaculture.
Keywords
microbial communities, biomarkers, aquaculture, El Niño–Southern
Oscillation (ENSO)
Picophytoplankton: harbingers of change in our coastal oceans
Download this Time Series Report
Citing this report:
Thomson P.G, Frampton D.M.F, Clementson L.A, Pattiaratchi C.B. Picophytoplankton: harbingers of change in our costal oceans. In Richardson A.J, Eriksen R, Moltmann T, Hodgson-Johnston I, Wallis J.R. (Eds). State and Trends of Australia’s Ocean Report. doi: 10.26198/5e16a9dd49e7e
doi: 10.26198/5e16a9dd49e7e


Citing the Report
Richardson A.J, Eriksen R, Moltmann T, Hodgson-Johnston I, Wallis J.R. (2020). State and Trends of Australia’s Ocean Report, Integrated Marine Observing System (IMOS).
The State and Trends of Australia's Ocean Report was supported by IMOS. IMOS gratefully acknowledges the additional support provided by the Commonwealth Scientific and Industrial Research Organisation (CSIRO).
The State and Trends of Australia's Ocean website is maintained by IMOS.

Australia’s Integrated Marine Observing System (IMOS) is enabled by the National Collaborative Research Infrastructure Strategy (NCRIS). It is operated by a consortium of institutions as an unincorporated joint venture, with the University of Tasmania as Lead Agent.
Disclaimer:
You accept all risks and responsibility for losses, damages, costs and other consequences resulting directly or indirectly from using this site and any information or material available from it. While the Integrated Marine Observing System (IMOS) has taken reasonable steps to ensure that the information on this website and related publication is correct, it provides no warranty or guarantee that information provided by the authors is accurate, complete or up-to-date. IMOS does not accept any responsibility or liability for any actions taken as a result of, or in reliance on, information on its website or publication. Users should check with the originating authors to confirm the accuracy of the information before taking any action in reliance on that information.
If you believe any information on this website or in the related publication is inaccurate, out of date or misleading, please bring it to our attention by contacting the authors directly or emailing us at IMOS@imos.org.au
Images and Information:
All information on this website remains the property of those who authored it. All images on this website are licensed through Adobe Stock, Shutterstock, or have permission from the original owner.